
不同基质配比下黄瓜根际细菌群落结构及多样性分析
2024-06-05 15:00陈宏昊唐小付聂圣贤林佳佳魏西刘婷任昊奎
热带作物学报 2024年3期
陈宏昊 唐小付 聂圣贤 林佳佳 魏西 刘婷 任昊奎

关键词:黄瓜;高通量测序;细菌群落结构;功能预测
黄瓜(Cucumber sativus L.)是我国重要的设施蔬菜[1],作为城市居民“菜篮子”中最主要的种类之一,具有较高的食用及营养价值特性,市场需求量大,在全国范围内广泛栽培。为了实现周年生产和季节性均衡供应,生产上一般采用日光温室、塑料拱棚、遮阳网等设施栽培,由于我国黄瓜设施栽培面积较大,复种指数高,常导致土壤板结,微生物群落平衡破坏[2-3]以及土传病害的发生,对黄瓜产量和品质造成不可避免的负面影响。相比之下,基质栽培能够有效预防和减少土传病害的发生[4-5],还能够一定程度上改善栽培作物的根系生长环境,提高产量和品质,因此是近年来新兴的栽培模式。
微生物是生态系统中功能活跃,开发潜力最大、最宝贵、最丰富的生物资源[6]。根际微生物与植物的生长密切相关,主要表现在微生物菌落结构、物种及多样性方面与植物的健康生长呈正向关联关系[7-9],尤其是一些根际细菌类群,具有较高的固氮、解磷、解钾和产植物生长素等[5]。大量研究表明,内生细菌及根际细菌种群可以定殖在植物根际或其他组织内[10],具有抑制病原菌生长,诱导植物增强抗病能力、促进生长等作用[11]。
无土基质栽培技术的实施与开展,改变了黄瓜在土壤中栽培的固定模式,越来越多的种植户选择其他基质进行栽培[12-13]。现阶段,基质栽培已经应用于沃柑[14]、番茄[15]、辣椒[16]、莴苣[17]、蔬菜[18]、芍药[19]等经济作物及植物的生产和种植。生产中,常用的商品基质主要成分草炭为不可再生资源,受环境和湿地保护的制约,其采挖已经受到限制[20]。目前,常见的复配基质材料有菇渣、椰糠、锯末、花生壳等,这些材料价格低廉,简单易得,可以通过改变材料种类和配比以获得不同作物的最佳配方。周方园等[5]研究发现,用基质栽培黄瓜可筛选出对黄瓜幼苗有显著促进作用的有益微生物,而胡云等[21]证明,基质与生物炭可提升黄瓜根际速效磷、速效钾、碱解氮等的质量比,从而提高黄瓜产量。
虽然前人对基质栽培黄瓜根际微生物多样性做了相关研究,但目前关于菇渣、锯末按不同比例复配对黄瓜根际菌落环境的影响方面研究鲜见报道。为此,本研究采用第三代测序技术比较分析了菇渣、锯末不同比例的复配基质对黄瓜植株根际细菌群落的影响,为菇渣、锯末复配的新型基质栽培模式应用提供依据。
1 材料与方法
1.1 材料
1.1.1 试验地概况 试验地位于广西壮族自治区南宁市西乡塘区广西大学农学院蔬菜试验基地(22°51.2′N,108°18′E)。
1.1.2 试验材料 供试黄瓜品种为津优186 号,购买自天津科润农业科技股份有限公司。
供试基质:菇渣(凤尾菇菇渣),购买自南宁市五塘食用菌生产有限公司;锯末(杉木锯末),购买自南宁市五塘木材加工有限公司;商品基质(泥炭∶蛭石∶珍珠岩=3∶1∶1)购买自长春益农赛世泥炭开发有限公司。
试剂盒:DNA 抽提试剂盒E.Z.N.A.? SoilDNA Kit 购买自美国Omega Bio-Tek 公司,建库试剂盒NEXTFLEX? Rapid DNA-Seq Kit 购买自美国Bioo Scientific 公司,测序试剂盒MiSeqReagent Kit v3 购买自美国Illumina 公司。
1.2 方法
1.2.1 黄瓜种植与产量测定 试验于2021 年7月至2022 年2 月在实验室和蔬菜基地进行。采用底袋直径和袋高均为40 cm 的无纺布袋进行基质袋装栽培。试验基质均用50%的多菌灵1000 倍液喷施消毒,然后覆盖塑料薄膜,在阳光下暴晒7 d。试验采用单因素隨机区组试验设计,共设4 种不同的栽培模式,每种栽培模式为一个试验处理,共4 个处理,每个处理设3 个重复,共12 个试验小区。各处理分别为:(1)CK 处理,商品基质对照;(2)T1 处理,复配基质(菇渣∶锯末=1∶3);(3)T2 处理,复配基质(菇渣∶锯末=1∶1);(4)T3 处理,复配基质(菇渣∶锯末=3∶1)。上述除栽培基质配比不同外,其他田间管理均相同。每个处理栽植黄瓜30 株,株距60 cm,行距80 cm。于9 月下旬进行穴盘育苗播种,10 月中旬定植,其他管理同日常,整个栽植过程中黄瓜植株未出现病虫害,12 月下旬在黄瓜进入收获期,按照小区对各处理的黄瓜依次分批采收,用常规称重法对黄瓜进行称重测产并记录。
1.2.2 样品采集 在黄瓜进入盛果期,每个处理选择10 株生长势一致的植株,采用“斗根法”[22]采集样品。取样时,先用75%酒精对手套和铁铲消毒,然后挖取植株根部,抖落根系外部基质,收集附着在植株根部的基质,装入无菌袋封装,放入冰盒,带回实验室。
1.2.3 样品总DNA 提取、PCR 扩增及高通量测序 使用E.Z.N.A.? Soil DNA Kit 试剂盒提取总DNA,使用NanoDrop 2000 超微量分光光度计(thermo fisher scientific, 美国)测定DNA 浓度和纯度,使用1%琼脂糖凝胶电泳检测总DNA 提取质量。PCR 扩增引物为799F(5-AACMGGATTAGATAC CCKG-3))和1193R(5-ACGTCATCCCCACCTTCC-3)),对样本细菌扩增V5-V7 可变区域进行PCR 扩增,其扩增参数为:95 ℃预变性3 min;95 ℃变性30 s,55 ℃退火30 s,72 ℃延伸30 s,循环27 次;72 ℃延伸10 min,10 ℃保存直至反应结束。PCR 扩增体系为:0.4 μLTaKaRa Taq(5 U/μL),4.0 μL 5×FastPfu Buffer,2.0 μL dNTPs(各2.5 mmol/L),0.8 μL(5 μmol/L)引物,0.2 μL 的BSA,1 μL DNA 模板(10 ng/μL),加入ddH2O 补足20 μL。对PCR 产物进行鉴定:即每个样本3 个PCR 重复,将3 个重复的PCR产物混合;使用2%琼脂糖凝胶电泳检测产物条带大小。Illumina Miseq 测序:使用2%琼脂糖凝胶回收PCR 产物,依次对回收产物进行纯化,再用2%琼脂糖凝胶进行电泳检测,并使用QuantusTMFluorometer(Promega, USA)对回收产物进行检测定量。使用NEXTFLEX? Rapid DNA-Seq Kit进行建库,使用MiSeq PE300 平台进行测序。
1.3 数据处理
试验数据采用Excel 2019 和SPSS 25.0 软件进行分析和处理, 用邓肯法进行显著性检验(P<0.05),平均数据以“平均数±标准差(SD)”表示。利用上海美吉生物医药科技有限公司I-Sanger 云数据分析平台对测序数据进行在线处理,获得样本稀释曲线图、OUT 聚类分析表格、Alpha 数据表格、细菌群落占比图、Venn 图及PICRUSt 功能预测数据等。
2 结果与分析
2.1 不同基质配比对黄瓜产量的影响
由表1 可知,菇渣、锯末按不同比例复配组成的复配基质(T1、T2、T3 处理)栽培黄瓜产量均显著高于商品基质(CK)栽培,其中T2 处理黄瓜产量最高,达48 572.40 kg/hm2;CK 处理产量最低,为39 762.70 kg/hm2。产量表现为T2>T3>T1>CK 处理,说明较CK 处理栽培相比,通过菇渣、锯末复配组成的基质栽培黄瓜可显著提高黄瓜产量,其中T2 处理产量最高。
2.2 不同基质配比下黄瓜植株根际细菌OUT聚类分析
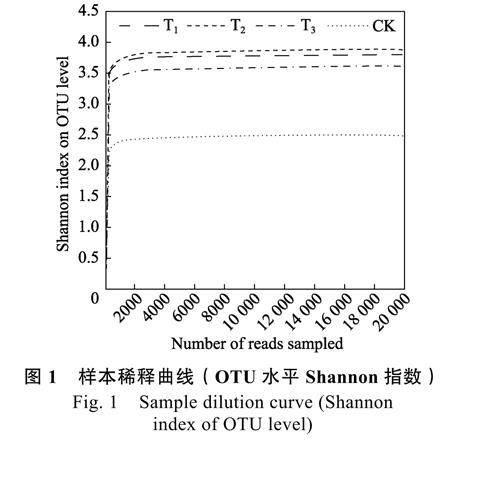
由图1 可知,稀释曲线随着样本测序数量的增大,逐渐变得平缓,当样本测序数量达到20000时,曲线不再增长,表明测序结果可以真实反映黄瓜根际细菌群落结构。
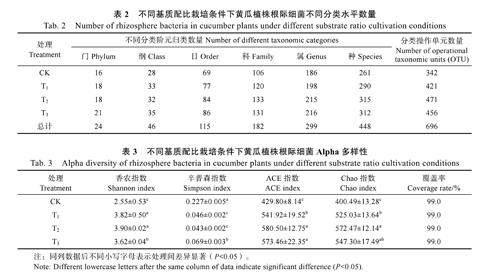
由表2 可知,不同栽培基质条件下黄瓜根际细菌菌落的组成存在较大差异,与CK 相比,T1、T2 和T3 处理均不同程度地提高了黄瓜根际细菌种群的多样性。
2.3 不同基质配比下黄瓜植株根际细菌Alpha多样性分析
在生物信息云平台进一步分析不同基质配比下黄瓜根际细菌的多样性指数和丰富度。由表3可知,各样本分析测序覆盖率均在99%以上,表明结果可以代表根际微生物种群的真实情况。T1、T2 和T3 处理的根际细菌Shannon 指数、ACE 指数和Chao 指数均显著高于CK 处理,说明T1、T2 和T3 处理栽培条件下,黄瓜植株根际微生物多样性和丰富度高于CK,且Simpson 指数均显著低于CK,表明商品基质栽培条件下黄瓜植株根际微生物多样性较低。
香农指数Shannon 和辛普森指数Simpson 是微生物多样性指标。香农指数其值越大,物种多样性越大,辛普森指数其值越大,物种越集中,多样性越低;Chao[23]指数和ACE[24]指数是微生物物种丰富度指标,其值越大,物种越丰富[25]。
2.4 群落组成分析
2.4.1 不同处理黄瓜植株根际细菌优势群落门分类水平 由图2 可知,门分类水平,不同基质配比下黄瓜植株根际细菌中,相对丰度占比大于1%的为优势细菌门,变形菌门(Proteobacteria)是4种栽培模式下共有的优势细菌门类,其相对丰度在各处理中占比介于72.45%~96.35%之间。
在CK 处理中,黄瓜植株根系优势细菌门的相对丰度占比大小依次为变形菌门(Proteobacteria,96.35%)、放线菌门(Actinobacteriota,1.43%),其他门类相对丰度总占比为1.49%。
在T1 处理中,黄瓜植株根系优势细菌门的相对丰度占比大小依次为变形菌门(Proteobacteria,72.45%)、拟杆菌门(Bacteroidota,25.14%),其他门类相对丰度总占比为1.48%。
在T2 处理中,黄瓜植株根系优势细菌门的相对丰度占比大小依次为变形菌门(Proteobacteria,80.78%)、拟杆菌门(Bacteroidota,16.51%),其他门类相对丰度总占比为1.73%。
在T3 处理中,黄瓜植株根系优势细菌门的相对丰度占比大小依次为变形菌门(Proteobacteria,84.04%)、拟杆菌门(Bacteroidota,12.49%)、放线菌门(Actinobacteriota,1.25%),其他门类相对丰度总占比为2.22%。
與CK 相比,虽然不同的基质配比栽培条件下均不同程度地降低了变形菌门(Proteobacteria)细菌丰度占比,但亦不同程度地增加了拟杆菌门( Bacteroidota ) 细菌丰度占比, 且拟杆菌门(Bacteroidota)是有机基质栽培特有的优势细菌门类。
2.4.2 不同处理黄瓜株根际细菌优势菌落属分类水平 如图3 所示,属分类水平,不同基质配比下黄瓜植株根际细菌中,相对丰度占比大于1%的为优势细菌属。其中在CK、T1、T2、T3 处理中,优势细菌属分类水平数量分别为12、15、15和14 个。
在CK 处理中,黄瓜植株根系优势细菌属的相对丰度占比大小顺序依次为罗丹诺杆菌属( Rhodanobacter , 46.40% )、不粘柄菌属(Asticcacaulis,14.77%)、包特菌属(Bordetella,12.02% )、伯克霍尔德氏菌属( Burkholderia-Caballeronia-Paraburkholderia,2.60%)、卡斯特兰尼氏菌属(Castellaniella,2.17%)、利姆诺杆菌属( Limnobacter , 1.87% )、( unclassified_c__Gammaproteobacteria,1.78%)、Noviherbaspirillu(1.63%)、申氏杆菌属(Shinella,1.62%)、德沃斯氏菌属( Devosia , 1.27% )、朱氏杆菌属(Chujaibacter,1.16%)、(unclassified_f__Comamonadaceae,1.05%),其他相对丰度总占比为8.47%。
在T1 处理中,黄瓜植株根系优势细菌属的相对丰度占比大小顺序依次为黄杆菌属(Flavobacterium,24.90%)、不粘柄菌属(Asticcacaulis,9.69% )、噬酸菌属( Acidovorax , 8.74% )、(unclassified_f__Comamonadaceae,6.68%)、伯克霍尔德氏菌属( Burkholderia-Caballeronia-Paraburkholderia , 4.63% )、罗丹诺杆菌属(Rhodanobacter,4.54%)、包特菌属(Bordetella,4.22%)、新鞘氨醇杆菌属(Novosphingobium,4.17%)、异样根瘤菌属( Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium,3.95%)、申氏杆菌属( Shinella , 3.67% )、慢生根瘤菌属( Bradyrhizobium, 2.52%)、草螺菌属(Herbaspirillum,2.23%)、德沃斯氏菌属(Devosia,2.05% )、( Dokdonella , 1.47% )、短波单胞菌(Brevundimonas,1.07%),其他相对丰度总占比为12.89%。
在T2 处理中,黄瓜植株根系优势细菌属的丰度占比大小顺序依次为黄杆菌属(Flavobacterium,15.89%)、不粘柄菌属(Asticcacaulis,14.74%)、伯克霍尔德氏菌属(Burkholderia-Caballeronia-Paraburkholderia,8.56%)、申氏杆菌属(Shinella,8.15%)、新鞘氨醇杆菌属(Novosphingobium,7.24%)、罗丹诺杆菌属(Rhodanobacter,6.5%)、噬酸菌属(Acidovorax,4.54%)、异样根瘤菌属( Allorhizobium-Neorhizobium-Pararhizobium-Rhizobium,3.65%)、(unclassified_f__Comamonadaceae,3.20%)、包特菌属(Bordetella,2.65%)、热单胞菌属( Thermomonas, 2.48%)、鞘氨醇盒菌属(Sphingopyxis,2.34%)、草螺菌属(Herbaspirillum,1.73%)、德沃斯氏菌属(Devosia,1.71%)、卡斯特兰尼氏菌属(Castellaniella,1.02%),其他相对丰度总占比为13.24%。
在T3 处理中,黄瓜植株根系优势细菌属的丰度占比大小顺序依次为罗丹诺杆菌属(Rhodanobacter,16.49%)、不粘柄菌属(Asticcacaulis,15.79%)、申氏杆菌属(Shinella,15.40%)、黄杆菌属(Flavobacterium,12.23%)、伯克霍尔德氏菌属(Burkholderia-Caballeronia-Paraburkholderia,4.21%)、新鞘氨醇杆菌属(Novosphingobium,3.89% )、异样根瘤菌属( Allorhizobium-NeorhizobiumPararhizobium-Rhizobium,3.37%)、噬酸菌属(Acidovorax , 2.28%)、德沃斯氏菌属(Devosia,2.11%)、草螺菌属(Herbaspirillum,1.97%)、热单胞菌属(Thermomonas,1.97%)、(unclassified_f__ Comamonadaceae,1.89%)、鞘氨醇盒菌属(Sphingopyxis,1.71%)、(Noviherbaspirillum,1.07%), 其他相对丰度总占比为12.14%。
其中,unclassified_c__Gammaproteobacteria、利姆诺杆菌属( Limnobacter ) 和朱氏杆菌属(Chujaibacter)是CK 处理条件下,黄瓜植株根系特有的优势细菌属;与CK 处理相比,黄杆菌属( Flavobacterium )、新鞘氨醇杆菌属(Novosphingobium)、异样根瘤菌属(Allorhizobium-NeorhizobiumPararhizobium-Rhizobium)、噬酸菌属Acidovorax 和草螺菌属(Herbaspirillum)是复配基质栽培(T1、T2、T3 处理)条件下,黄瓜植株根际特有的优势细菌属;伯克霍尔德氏菌属(Burkholderia-Caballeronia-Paraburkholderia)、罗丹诺杆菌属(Rhodanobacter)、不粘柄菌属( Asticcacaulis )、申氏杆菌属( Shinella ) 和unclassified_f__Comamonadaceae、德沃斯氏菌属(Devosia)是商品基质栽培和菇渣、锯末复配基质栽培条件下,黄瓜植株根际共有的优势细菌属。
综上所述,不同的基质配比栽培黄瓜,在不同程度上改变黄瓜植株根际细菌属分类水平的群落组成,同时也改变了优势细菌菌落组成的丰度占比。与CK 相比,菇渣、锯末复配基质栽培有利于黄瓜根际形成更为多样与均衡的细菌群落结构。
2.5 物种Venn 分析
由图4 分析结果可知,在属分类水平,CK、T1、T2、T3 处理中,黄瓜植株根际细菌属分类水平数量分别为165、192、198 和200 个;4 种栽培模式中,黄瓜植株根际细菌共有的菌属为102个;T1、T2、T3 与CK 处理共有的优势根际细菌属数量分别为14、6 和7 个。而T1、T2、T3 和CK 处理黄瓜植株根际中,特有的根际细菌属的数量分别为18、14、23 和14 个。
由图5 分析结果可知,在种分类水平,CK、T1、T2、T3 处理中,黄瓜植株根际细菌种分类水平数量分别为226、279、294 和286 个;4 种栽培模式中,黄瓜植株根际细菌共有的菌种为130个;T1、T2、T3、CK 处理共有的优势根际细菌种数量分别为14、10、7 个。而T1、T2、T3、CK处理黄瓜植株根际中,特有的根际细菌种的数量分别为35、27、34、28 个。
表明无论是属或种分类水平,与CK 相比,通过菇渣、锯末复配基质栽培均可以提高黄瓜根际细菌的数量水平,同时也有利于提升黄瓜植株根际特有的优势细菌数量。
2.6 PICRUSt 功能预测
为了进一步研究不同基质配比栽培条件下黄瓜植株根际细菌的功能,本研究利用高通量测序技术和KEGG(kyoto encyclopedia of genes andgenomes)数据库进行比对发现,黄瓜植株根际细菌在一级功能层共有6 类生物代谢通路(表4),分别为: 代谢(metabolism )、环境信息处理(environmental information processing)、遗传信息处理(genetic information processing)、细胞过程(cellular processes)、人类疾病(human diseases)和有機系统(organismal systems)。其中,代谢、环境信息处理和遗传信息处理为主要一级功能,占比分别为62.74%~64.24%、15.64%~17.43%和10.57%~11.07%。不同基质配比栽培条件下黄瓜植株根际细菌基因一级功能层预测基因种类无显著差异,但基因拷贝数占比存在差异。
COG(clusters of orthologous groups)功能分类如图6 所示,黄瓜植株根际细菌群落功能组成相对丰度占比前10 类别分别为:未知功能(S,11.06%~11.64%)、氨基酸转运与代谢(E,7.99%~8.19%)、一般功能预测(R,7.92%~8.17%)、细胞壁/膜/包膜生物发生(M,6.80%~7.09%)、转录(K,6.61%~6.81%)、无机离子运输与代谢(P,6.11%~6.61%)、能量生产和转换(C, 6.32%~6.60%)、信号转导机制(T,5.60%~6.45%)、碳水化合物运输和代谢(G,5.24%~6.06%)以及翻译、核糖体结构和生物发生(J,5.15%~5.43%)。各处理中,黄瓜植株根际细菌群落功能组成相似。与CK 相比,菇渣、锯末复配基质栽培提高了一般功能预测、氨基酸转运与代谢、转录、无机离子运输与代谢、信号转导机制、碳水化合物运输和代谢功能丰度,降低了能量生产和转换、细胞壁/膜/包膜生物发生、翻译、核糖体结构和生物发生等功能丰度,但不同基质配比栽培条件下各处理间差异不显著。表明不同基质配比对黄瓜植株根际细菌功能丰度占比影响较小。通过对KEGG数据库和COG 数据库检测到大多数根际细菌显示出有益的功能。
3 讨论
本研究中,基于黄瓜植株根际细菌群落结构组成和产量而言,菇渣和锯末复配基质可替代常规商品基质栽培黄瓜,T2 处理(菇渣∶锯末=1∶1)表现最佳,为新型基质栽培模式的应用提供理论参考依据。
基质栽培相较于传统的土壤栽培具有节水、保墒、增产及减轻环境污染,降低生产成本的作用,能够有效改善作物生长的根际环境,提高作物的产量和品质。根际微生物群落的形成与作物、土壤有着密切的关系,而根际微生物在作物对养分的吸收、病原物的防卫,抗逆性等方面发挥着至关重要的作用[26]。生产上通过间作制度、生物化肥、基质栽培等方式改善作物根际微生物群落组成和功能,并通过对根际微生物群落的调控来增加农作物的产量。已有研究表明,作物根际微生物或内生微生物群落结构和多样性越丰富时,植株对抗病原菌的综合能力就越强[27-28]。本研究发现,菇渣、锯末复配组成的基质栽培条件下Chao 指数和Shannon 指数均显著高于商品基质栽培,在菇渣、锯末复配基质栽培条件下,检测到黄瓜植株根际富集一些有益功能菌属如:黄杆菌属( Flavobacterium )、新鞘氨醇杆菌属(Novosphingobium)和草螺菌属(Herbaspirillum)等共有优势功能菌属。有研究发现,黄杆菌属(Flavobacterium)细菌具有脱氮除磷功能的好氧反硝化微生物[29] , 新鞘氨醇杆菌属(Novosphingobium)是一种与碳循环相关的具有降解木质素功能的微生物[30-31] , 草螺菌属(Herbaspirillum)是一种与根系相关的固氮细菌,可利用根系分泌物互利共生,其主要通过固氮、溶磷和分泌铁载体等方式促进植物生长[32]。可见,这些特有优势功能菌属的富集改善了黄瓜植株根际微生态环境,帮助植株适应养分不足的胁迫环境。
前人研究发现,基质栽培中,不同的基质配比能不同程度地改变沃柑大苗根际细菌的多样性和丰富度[14]。本研究亦发现,菇渣、锯末不同的基质配比调节了黄瓜植株根际优势细菌不同(门、属)分类水平的组成比例,不同程度地改变了黄瓜植株根际细菌的多样性和丰富度。杨海俊[33]研究发现,用粗砂、菇渣、锯末等作为复配基质材料,理化特性及变化满足作物生长需求,明显改善了作物根区的环境,提高了作物产量和品质,与本研究结果有相似之处,本研究發现,拟杆菌门(Bacteroidota)是菇渣、锯末复配基质栽培共有的优势细菌门类,能够有效降解有机物质,加快栽培基质中有机物料的分解与吸收,改善了黄瓜植株根区微生态环境,且产量也高于商品基质栽培。良好的栽培基质能为作物的生长提供良好的根际微生态环境,栽培基质原料与配比的不同不仅会影响基质理化性状,也会影响基质微生物的数量、种类及酶活性,进而影响植物的生长[34]。因此,研究菇渣、锯末不同的配比下黄瓜根际细菌群落结构的分析,对黄瓜生产及新的栽培模式具有重要意义。
近年来,利用高通量测序技术在细菌多样性及丰富度方面研究较多[35-38]。对细菌功能研究相对较少,本研究在Mi Seq 高通量测序结果的基础上,将KEGG 数据库和COG 数据库进行比对发现,不同的基质配比栽培黄瓜不仅改变了黄瓜植株根际细菌群落组成,而且影响植株根际细菌的代谢功能,但不同处理之间对黄瓜植株根际细菌群落整体功能影响甚微。今后关于细菌功能的研究及有益功能菌如何对作物的生长产生影响,有待进一步深入探究。
猜你喜欢
中老年保健(2021年5期)2021-08-24
今日农业(2020年16期)2020-12-14
小学生学习指导(低年级)(2020年10期)2020-11-09
作文大王·低年级(2019年11期)2019-12-02
小学生导刊(2017年19期)2017-07-19
中国医药导报(2017年9期)2017-05-11
中国中药杂志(2016年24期)2017-04-18
中国中药杂志(2017年4期)2017-03-28
中国中药杂志(2017年3期)2017-03-20
科技创新导报(2016年28期)2017-03-14
