
天平的两端:活化性和抑制性Fcγ受体
2016-04-10 07:40黄林芳廖湘平郑文洁
中华临床免疫和变态反应杂志 2016年1期
黄林芳,廖湘平,雷 丽,张 烜,郑文洁#
(南华大学附属郴州市第一人民医院风湿免疫科,湖南郴州 423000)
与免疫球蛋白(immunoglobulin,Ig)G的Fc片段结合受体(Fcγ receptors,FcγRs)表达在多种免疫细胞,但在不同的细胞类型上FcγRs的表达各异。FcγRs家族成员主要与IgG有低亲和力,因此只结合于免疫复合物,从而避免了IgG在没有抗原刺激时活化免疫细胞[1],当抗原抗体复合物偶联活化性Fcγ受体时引起免疫受体酪氨酸活化基序(immunoreceptor tyrosine-based activation motif,ITAM)磷酸化,继而活化下游信号分子SYK,启动信号级联;当与抑制性Fcγ受体(FcγRⅡB)偶联时,对下游目标去磷酸化,进而抑制活化性信号的级联[2-3]。FcγRⅡB还能通过与树突状细胞相关性C型植物血凝素-1(Dectin-1)的偶联阻断补体介导的炎性作用[4]。活化性FcγRs与FcγRⅡB的相对关系-被称为激活与抑制受体比例(activating to inhibitory,A:I),当抑制性Fcγ受体不能抗衡活化性Fcγ受体时,机体的平衡状态被扰乱,将导致免疫病的发生[1-2]。本文将从FcγRⅡB及其与活化性Fcγ受体之间比例的失调和不同免疫病的关系进行综述。
Fcγ受体家族
根据与IgG的亲和力及信号通路的不同,Fcγ受体分为高亲和力的FcγⅠ、低亲和力的FcγⅡA、FcγⅡB、FcγⅡC、FcγⅢA、FcγⅢB及FcγⅣ,其中FcγⅢ、FcγⅣ仅表达于老鼠,而FcγⅡB是唯一的抑制性Fcγ受体。单核巨噬细胞、树突状细胞、嗜碱性粒细胞及肥大细胞上既有活化性又有抑制性Fcγ受体的表达,两者维持一定的平衡关系。自然杀伤细胞只表达活化性受体FcγⅢ,而B细胞只表达抑制性受体FcγRⅡB(表1)[3]。
FcγRⅡB与不同细胞表面活化性受体的相互作用
B细胞
B细胞的活化是通过抑制性受体FcγRⅡB和活化性受体CD19及BCR之间的相互作用而调节的[5]。偶联于BCR时,FcγRⅡB胞质区ITIM结构域的酪氨酸被磷酸化,继而招募并激活含SH2结构域的酪氨酸磷酸酶-1(SHP1)、酪氨酸磷酸酶-2(SHIP2),使ITIM通路中3,4,5三磷酸酯酰基醇(PIP3)去磷酸化,阻断布鲁顿酪氨酸激酶(Btk)等在细胞表面的聚集,降低细胞内钙离子水平,从而抑制活化性受体可能引起的持续性的B细胞活化效应[5-6]。有研究发现,IC是通过偶联FcγRⅡB抑制TLR9的异常信号,从而降低B细胞的活化、减少相关自身抗体的产生[7]。XmAb5871是针对FcγRIIB的靶向治疗药物,通过交联B细胞表面受体及FcγRIIB,以达到抑制B细胞的异常活化及功能的作用,通过利用类风湿关节炎(rheumatoid arthritis,RA)患者的外周血单个核细胞(peripheral blood mononuclear cell,PBMC)进行XmAb5871刺激的体内及体外试验发现,FcγRIIB可以抑制B细胞产生的体液免疫反应[8]。因此,当B细胞上FcγRⅡB的表达降低时,将不能抑制活化性受体B细胞受体诱导的B细胞的异常活化,容易诱发B细胞相关的自身免疫病,如系统性红斑狼疮(systemic lupus erythematosus,SLE)、RA等。
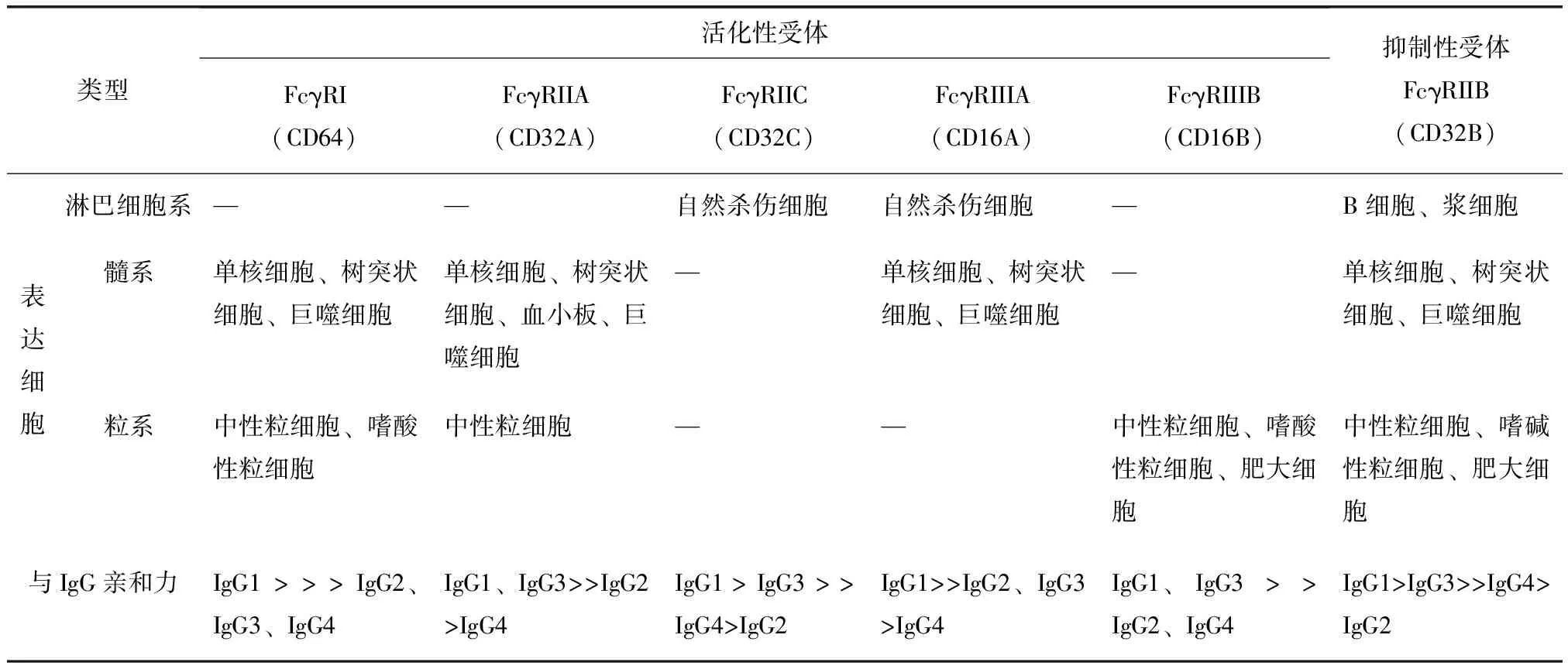
表1 人类活化性受体与抑制性受体的表达分布及与IgG亚型亲和力比较
FcγR:Fc片段结合受体
单核巨噬细胞
FcγRⅡB通过偶联活化性Fcγ受体,抑制其介导的吞噬作用和相关细胞因子[肿瘤坏死因子(tumor necrosis factor,TNF)-α、白细胞介素(interleukin,IL)-6、IL-1a)]的释放,同时抑制Toll样受体(Toll-like receptors,TLR)4介导的活化[2,9]。有研究表明,来源于FcγRⅡB敲除小鼠的单核巨噬细胞比正常小鼠的更容易活化,且这些小鼠更容易出现免疫复合物诱导的肺泡炎、关节炎和败血症,但是,由于这些小鼠的巨噬细胞清除细菌的能力增强,它们能够更强地抵制肺炎链球菌的感染,因此,诱导巨噬细胞过表达FcγRⅡB的小鼠,在感染肺炎链球菌后死亡率是增加的[2]。有研究通过构建大鼠FcγRⅡB慢病毒诱导表达载体,经检测巨噬细胞上FcγRⅡB的表达后发现,巨噬细胞的吞噬和趋化功能与FcγRⅡB的表达呈负相关[9]。因此,FcγRⅡB表达过高或过低都会导致疾病的发生,维持FcγRⅡB与活化性Fcγ受体之间的平衡关系非常重要[10]。
树突状细胞
FcγRs在增强树突状细胞(dendritic cells,DC)的抗原提成作用方面起着关键作用,尽管抑制性受体FcγRⅡB和活化性Fcγ受体都能介导免疫复合物来源的抗原提成,但从它们活化DC及刺激MHCⅠ类分子交叉提呈的能力方面显示,活化性Fcγ受体在促进抗原提呈时起主要作用[10]。相关研究也表明,在DC对T细胞的活化作用中,FcγRⅡB发挥了主要作用,FcγRⅡB表达缺失的DC细胞,其活化初始T细胞的能力显著增强,CD8+T细胞的表达明显增加[11]。
中性粒细胞
研究表明,TNFα能上调中性粒细胞上活化性受体FcγRIIA的表达,从而增强免疫复合物诱导的中性粒细胞的活化。TNFα抑制剂英夫利昔单抗(infliximab)在降低FcγRIIA的同时增加FcγRⅡB的表达,修复活化性Fcγ受体与抑制性Fcγ受体的比例失调,增加免疫复合物介导的中性粒细胞活化的阈值,显著减轻RA患者的炎性反应[12]。
FcγR在不同自身免疫性疾病的表现
FcγR存在于不同细胞表面,通过调节各细胞反应的激活阈,进而抑制IgG介导的免疫效应,维持机体的外周免疫耐受平衡。活化性Fcγ受体表达增加或抑制性Fcγ受体表达降低都会导致多种免疫性疾病的发生[13]。
系统性红斑狼疮
B细胞的功能紊乱及免疫复合物的清除障碍与SLE的发病密切相关,FcγRⅡB作为B细胞的一个关键调节器,在SLE的发病中起着重要作用[14]。动物实验表明,FcγRⅡB敲除的C57BL/6小鼠自发产生高丙种球蛋白、自身抗体和肾小球肾炎等免疫复合物介导的狼疮样自身免疫病的表型,纠正FcγRⅡB的表达后能改善患病小鼠的病情,明显减少小鼠SLE发病率[2,15]。另有研究发现,活动性SLE患者记忆B细胞和浆细胞表面FcγRⅡB的表达明显低于其初始B细胞及正常健康对照的记忆B/浆细胞[16]。SLE患者单核细胞上活化性受体FcγⅠ的表达高于健康对照,尤以伴有狼疮肾炎者明显,且FcγⅠ表达水平与患者肾功能损害(血肌酐、尿素氮、微量白蛋白/肌酐水平)和超敏C反应蛋白水平呈正相关,而与补体C3水平呈负相关;高表达FcγⅠ的单核细胞分泌炎性趋化因子单核细胞趋化因子(monocyte chemotactic factor,MCP)-1增加;激素(>40mg/d)治疗后,FcγⅠ的水平减低[17]。Jeon等[18]发现,朝鲜SLE患者FcγRIIB(rs12117530)的基因多态性与疾病的严重程度和临床表现相关。国内学者也发现,SLE患者纯合子FcγRIIB-I232T突变体的原代B细胞,不能阻止B细胞受体与CD19的突触共定位,导致下游信号传导分子对PI3K的异常调节,促使了B淋巴细胞的异常活化[5]。
类风湿关节炎
胶原性关节炎(collagen induced arthritis,CIA)小鼠模型中,FcγRⅡB的缺乏增加胶原特异性的抗体滴度和疾病的严重程度,并且 B 细胞FcγRⅡB转基因鼠的疾病严重程度和抗体滴度都降低,同时FcγRⅡB通过抑制基质金属蛋白酶的产生限制关节软骨的破坏[2]。Catalán等[19]证实,RA患者记忆B细胞及浆母细胞表面FcγRⅡB表达降低,且与高浓度自身抗体具有相关性。Magnusson等[20]发现,RA患者滑膜组织中巨噬细胞FcγRⅡB及活化性FcγI、FcγII、FcγIII受体的表达均增加,且局部关节腔注射抗炎药物后,滑膜中FcγRⅠ表达降低,提示FcγR在RA滑膜炎症发挥中心作用。另有研究发现,RA患者单核细胞的活化性Fcγ受体FcγRⅠ和FcγRII a增加,而FcγRⅡB表达与健康人无明显差别;活化性与抑制性Fcγ受体失衡,可增加单核/巨噬细胞相关炎性细胞因子分泌,加重RA的发病,而IL-4联合IL-10刺激则能增加FcγRⅡB表达,减少炎症细胞因子释放[21]。Chen等[22]的研究表明,FcγRⅡB 187异亮氨酸等位基因的多态性与RA患者抗环瓜氨酸肽抗体(抗cyclic citrullinated peptide抗体,抗CCP抗体)、类风湿因子(rheumatoid factor,RF)的产生及疾病的严重程度密切相关,且纯合子发病早,多见于出现骨侵蚀的男性患者。
原发性干燥综合征
原发性干燥综合征(primary Sjögren’s syndrome,pSS)患者活动期外周血记忆性B细胞FcγRⅡB的表达降低,且FcγRⅡB的表达与其活动指数pSS疾病活动度(Sjögren’s syndrome disease activity index,SSDAI)呈负相关,并与抗SSA抗体呈负相关[23]。因此,FcγRⅡB表达异常可能在pSS免疫发病机制中也起着重要作用。
银屑病性关节炎
银屑病关节炎(psoriatic arthritis,PsA)滑膜组织中巨噬细胞的数量增加,针对单核巨噬细胞的血浆吸附治疗有助于缓解其病情,表明单核巨噬细胞参与其发病[24]。有研究发现,PsA 患者单核细胞上FcγRⅠ的表达明显高于健康对照,且与病情活动指标[红细胞沉降率(erythrocyte sedimentation rate,ESR)、C反应蛋白(C reactive protein,CRP)、病情活动度评分(disease activity score 28,DAS28)和总IgG水平]呈正相关,提示纠正FcγRⅠ的异常表达可能利于治疗PsA[25]。
原发性血小板减少性紫癜
特发性血小板减少性紫癜(idiopathic thrombocytopenic purpura,ITP)患者FcγRⅠ的表达增加,用大剂量激素治疗ITP患者发现,治疗前单核细胞上FcγRⅡA/FcγRⅡB mRNA的比例高于健康对照,并伴随异常的吞噬能力,在大剂量激素治疗后发现,FcγRⅡB mRNA的表达水平明显增加,而FcγRⅡA、FcγRⅠ的表达水平及单核细胞的吞噬能力都下降,两者比例恢复正常[26]。同时,针对ITP患者幽门螺杆菌的消除治疗也有类似结果,FcγRⅡB的表达与幽门螺杆菌的表达呈负相关,幽门螺杆菌阳性的ITP患者FcγRⅡA/FcγRⅡB mRNA的比例高于阴性患者,治疗后单核细胞的异常吞噬能力降低,血小板数量恢复正常,活化性Fcγ受体与抑制性Fcγ受体之间比例恢复平衡[2,27]。综上,说明活化性Fcγ受体与抑制性Fcγ受体共同参与了ITP的发病,通过对他们的研究可能为ITP的治疗提供新的治疗靶点。
其他
桥本甲状腺炎:与健康人相比,桥本甲状腺炎患者外周血单核细胞表面FcγRⅢ的表达显著高于健康对照;而B细胞上FcγRⅡB的表达明显低于健康对照,提示桥本甲状腺炎患者存在活化性Fcγ受体与抑制性Fcγ受体平衡的紊乱[28]。
慢性炎性脱髓鞘性多发性神经根神经病:慢性炎性脱髓鞘性多发性神经根神经病(chronic inflammatory demyelinating polyneuropathy,CIDP)患者B细胞上FcγRⅡB的表达下降,单核细胞上FcγRⅡB的表达也低于健康对照,但活化性受体FcγRⅠ表达增加;人免疫球蛋白治疗可部分恢复以上的FcγR表达失衡,因此,通过平衡两者之间的表达水平将为治疗CIDP提供新的靶点[29]。
结论和展望
FcγRs家族在体内多种细胞上均有表达,通过不同的机制维持机体免疫平衡[30],在调节体液免疫、抗体介导的效应功能和细胞免疫反应中起着关键作用,在促炎症和抗炎症反应中的作用也越来越受到重视。大量研究证明,FcγRⅡB异常或其与活化性受体的比例失衡将导致多种疾病的发生,因此,加强对FcγRs的了解有助于探索疾病的易感性及发现更好的治疗策略。
[1]Espéli M,Smith KG,Clatworthy MR.FcγRIIB and autoimmunity[J].Immunol Rev,2016,269:194-211.
[2]Smith KG,Clatworthy MR.FcγRIIB in autoimmunity and infection:evolutionary and therapeutic implications[J].Nat Rev Immunol ,2010,10:328-343.
[3]Nimmerjahn F,Ravetch JV.Fcγ receptors as regulators of immune responses[J].Nat Rev Immunol,2008,8:34-47.
[4]Karsten CM,Pandey MK,Figge J,et al.Galactosylated IgG1 links FcγRIIB and Dectin-1 to block complement-mediated inflammation[J].Nat Med,2012,18:1401.
[5]Xu L,Li G,Wang J,et al.Through an ITIM-Independent Mechanism the FcγRIIB Blocks B Cell Activation by Disrupting the ColocalizedMicroclustering of the B Cell Receptor and CD19[J].J Immunol,2014,192:5179-5191.
[6]姚忻,闻玉梅.抑制性IgG受体 (FcγRIIB) 的免疫调节作用及意义[J].生命科学杂志,2009,21:49-52.
[7]Qian L,Chen W,Qin H,et al.Immune complex negatively regulates Toll-like receptor 9-mediated immune responses in B cells through the inhibitory Fc-gamma receptor IIb[J].Microbiol immunol,2015,59:142-151.
[8]Chu SY,Yeter K,Kotha R,et al.Suppression of Rheumatoid Arthritis B Cells by XmAb5871,an Anti-CD19 Antibody That Coengages B Cell Antigen Receptor Complex and Fcγ Receptor IIb Inhibitory Receptor[J].Arthritis Rheumatol,2014,66:1153-1164.
[9]郑修军,王琦,曹秀琴,等.上调巨噬细胞FcγRⅡB的表达可抑制其吞噬和趋化功能[J].细胞与分子免疫学杂志,2015:1022-1026.
[10] Guilliams M,Bruhns P,Saeys Y,et al.The function of Fc [gamma] receptors in dendritic cells and macrophages[J].Nat Rev Immunol,2014,14:94-108.
[11] van Montfoort N,’tHoen PA,Mangsbo SM,et al.Fcγ receptor IIb strongly regulates Fcγ receptor-facilitated T cell activation by dendritic cells[J].J Immunol,2012,189:92-101.
[12] Belostocki K,Pricop L,Redecha PB,et al.Infliximab treatment shifts the balance between stimulatory and inhibitory Fcγ receptor type II isoforms on neutrophils in patients with rheumatoid arthritis[J].Arthritis Rheum,2008,58:384-388.
[13] Ravetch JV,Lanier LL.Immune inhibitory receptors[J].Science,2000,290:84-89.
[14] 卢志明,陈国建,崔相法,等.外周血单个核细胞FcTRⅡb和血清C_1q抗体变化在SLE发病机制中的作用[J].山东医药,2005,45:1-2.
[15] McGaha TL,Sorrentino B,Ravetch JV.Restoration of tolerance in lupus by targeted inhibitory receptor expression[J].Science,2005,307:590-593.
[16] Su K,Yang H,Li X,et al.Expression profile of FcγRIIb on leukocytes and its dysregulation in systemic lupus erythematosus[J].J Immunol,2007,178:3272-3280.
[17] Li Y,Lee PY,Sobel ES,et al.Increased expression of FcgammaRICD64 on circulating monocytes parallels ongoing inflammation and nephritis in lupus[J].Arthritis Res Ther,2009,11:R6.
[18] Jeon JY,Kim KY,Kim BS,et al.FcγRIIB Gene Polymorphisms Are Associated with Disease Risk and Clinical Manifestations of Systemic Lupus Erythematosus in Koreans[J].Tohoku J Exp Med,2015,236:185-191.
[19] Catalán D,Aravena O,Sabugo F,et al.B cells from rheumatoid arthritis patients show important alterations in the expression of CD86 and FcgRIIb,which are modulated by anti-tumor necrosis factor therapy[J].Arthritis Res Ther,2010,12:R68.
[20] Magnusson SE,Engström M,Jacob U,et al.High synovial expression of the inhibitory FcγRIIb in rheumatoid arthritis[J].Arthritis Res Ther,2007,9:R51.
[21] Wijngaarden S,van de Winkel JG,Jacobs KM,et al.A shift in the balance of inhibitory and activating Fcγ receptors on monocytes toward the inhibitory Fcγ receptor IIb is associated with prevention of monocyte activation in rheumatoid arthritis[J].Arthritis Rheum,2004,50:3878-3887.
[22] Chen JY,Wang CM,Ma CC,et al.A transmembrane polymorphism in FcγRIIb (FCGR2B) is associated with the production of anti-cyclic citrullinated peptide autoantibodies in Taiwanese RA[J].Genes Immun,2008,9:680-688.
[23] 周士亮,车楠,陆智敏,等.原发性干燥综合征患者外周血 B 细胞FcγRⅡB的表达及临床意义[J].中华风湿病学杂志,2012,16:292-295.
[24] Kanekura T,Kawabata H,Maruyama I,et al.Treatment of psoriatic arthritis with granulocyte and monocyte adsorption apheresis[J].J Am Acad Dermatol,2004,50:242-246.
[25] Matt P,Lindqvist U,Kleinau S.Up-regulation of CD64-expressing monocytes with impaired FcγR function reflects disease activity in polyarticular psoriatic arthritis[J].Scand J Rheumatol,2015,44:464-473.
[26] Liu XG,Ma SH,Sun JZ,et al.High-dose dexamethasone shifts the balance of stimulatory and inhibitory Fcγ receptors on monocytes in patients with primary immune thrombocytopenia[J].Blood,2011,117:2061-2069.
[27] Asahi A,Nishimoto T,Okazaki Y,et al.Helicobacter pylori eradication shifts monocyte Fcγ receptor balance toward inhibitory FcγRIIB in immune thrombocytopenic purpurapatients[J].J Clin Invest,2008,118:2939.
[28] Liu Y,Liu M,Zhang Y,et al.The expression of Fcγ receptors in Hashimoto’s thyroiditis[J].Cell Immunol,2015,294:33-38.
[29] Quast I,Cueni F,Nimmerjahn F,et al.Deregulated Fcγ receptor expression in patients with CIDP[J].Neurol Neuroimmunol Neuroinflamm,2015,2:e148.
[30] Li F,Smith P,Ravetch JV.Inhibitory Fcγ receptor is required for the maintenance of tolerance through distinct mechanisms[J].J Immunol,2014,192:3021-3028.
猜你喜欢
中小企业管理与科技(2022年19期)2022-12-22
武汉大学学报(医学版)(2022年2期)2022-11-23
九江学院学报(自然科学版)(2022年2期)2022-07-02
食品与发酵工业(2022年9期)2022-05-17
天津医科大学学报(2021年1期)2021-01-26
世界最新医学信息文摘(2020年11期)2020-12-25
中成药(2019年12期)2020-01-04
中国化工贸易·下旬刊(2019年6期)2019-10-21
浙江医学(2019年12期)2019-07-10
科学与财富(2016年38期)2017-05-23
